
高等学校生物/生物I/遺伝情報とDNA
高等学校生物 > 生物I > 遺伝
DNAの構造
[編集]


DNA(デオキシリボ核酸、英: deoxyribonucleic acid)の構造は、ヌクレオチド (nucleotide) と呼ばれる構成単位をもち、ヌクレオチドはリン酸と糖と塩基の化合物である。ヌクレオチドの糖はデオキシリボース(deoxyribose) である。DNAでは、ヌクレオチドがいくつも結合して、二重らせん構造をつくっている。
塩基には4種類あり、アデニン(adenine)、チミン(thymine)、シトシン(cytosine)、グアニン(guanine)という4種類の塩基である。ヌクレオチド一個に、4種の塩基のうち、どれか一個が、ふくまれる。
生殖細胞では、減数分裂で染色体が半分になることから、遺伝子の正体とは、どうやら染色体に含まれている物質であろう、という事がモーガンなどの1913年ごろのショウジョウバエの遺伝の研究によって、突き止められていた。
遺伝子に含まれる物質にはタンパク質や核酸(かくさん)など、さまざまな物質がある。どの物質こそが遺伝子の正体なのかを突き止める必要があった。核酸の発見は、1869年ごろ、スイスの生化学者ミーシャーによって、膿(うみ)から取り出した細胞の核に、リン酸をふくんだ物質があることが発見され、この物質はタンパク質とは異なることが調べられた。ミーシャ-の発見したのが核酸である。この当時では、まだ核酸が遺伝子の正体だとは気づかれていなかった。なお、膿は、白血球を多くふくむ。
1949年、オーストリアのエルヴィン・シャルガフは、 いろいろな生物の持つDNAを抽出して調べ、どの生物でもアデニン(A)とチミン(T)とは量が等しく1:1であり、グアニン(G)とシトシン(C)とは量が等しく1:1であることを発見した。
- A:T = 1:1 、 G:C = 1:1
このことから、シャルガフは、アデニンはチミンと結合する性質があり、グアニンはシトシンと結合する性質があると考えた。 DNAの、このような、アデニン(A)とチミン(T)とが等量で結合する性質があること、グアニンとシトシンも等量で結合する性質があることを、まとめて、相補性(そうほせい)という。
1953年、アメリカのジェームズ・ワトソンとイギリスのフランシス・クリックは、 シャルガフの塩基組成の研究や、イギリスのモーリス・ウィルキンスのX線回折の研究をもとにして、研究を行った。そしてワトソンとクリックは、DNAが二重らせん構造であることを発見した。 これによると、2本のヌクレオチド鎖が、アデニンとチミン、グアニンとシトシンで対合し、柱状になり、それがらせん状にねじれている。
-
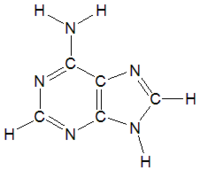 アデニン(A)
アデニン(A) -
 チミン(T)
チミン(T) -
 グアニン(G)
グアニン(G) -
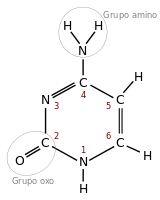 シトシン(C)
シトシン(C)






二重らせん上のアデニンAとチミンTなど、らせんで対になった塩基どうしの結合は、水素結合(すいそ けつごう)という、水素を仲立ちとした弱い結合をしている。塩基上の水素原子が、向かいあった塩基の窒素原子や酸素原子などと、弱く結合するのが、DNAの場合での水素結合である。
なお水素結合が見られるのは生物だけに限らず、一般の化学物質などでも多く見られる。たとえば水分子の安定性でも、水素結合が関わっている。
DNAの場合の水素結合では、アデニンはチミンの塩基対では、塩基上の2箇所で水素結合をする。シトシンとグアニンの塩基対では、塩基上の3箇所で水素結合をする。
二重らせんの幅は2.0nmで、らせん1回転(1ピッチ)の長さは3.4nm、らせん1回転中に10対のヌクレオチド対がある。
DNAの働き
[編集]

DNAの働きには、主にタンパク質の設計図となることと、遺伝情報を子孫に伝えることがある。
DNAの遺伝子の働きかたを決める要因は、塩基の並び方で決定される。この塩基の並び方で、細胞で合成されるタンパク質が異なるため、DNAはタンパク質の設計図となっている。このため、DNAの塩基の並び方が異なると、遺伝情報も異なる。病気などの例外をのぞけば、ある生体で合成されたタンパク質、たとえば皮膚のタンパク質のコラーゲンや、骨のタンパク質や、筋肉のタンパク質のミオシンなど、どのタンパク質も、その生体のDNAの情報をもとに合成されたタンパク質である。
DNAは、細胞核の中で、RNA(アールエヌエー)というタンパク質合成用の塩基配列の物質をつくる。RNAの情報は、DNAの情報を元にしている。RNAは、核の外に出ていきリボソームと結合し、消化器官で食品のタンパク質から分解・吸収したアミノ酸を材料にして、 RNAの塩基配列に従ってアミノ酸をつなぎかえることで、タンパク質を作っている。
タンパク質の構造は、アミノ酸がいくつも結合した構造である。したがって、タンパク質を構成するアミノ酸の順序などの配列や、アミノ酸の数などによって、タンパク質の性質が異なる。なお、アミノ酸どうしの化学結合をペプチド結合という。
- (※ タンパク質の合成の仕組みについて、くわしくは、単元『高等学校理科 生物基礎/遺伝情報とタンパク質の合成』などの章で説明する。)
DNAは、受精卵の時から、細胞分裂の際は、必ず複製されている。 DNAは配偶子形成の際半分になり、配偶子が受精すると合わさって元に戻る。 こうしてDNAは遺伝情報を子孫に伝えている。
- (※ 生殖細胞とDNAの分配の仕組みについて、くわしくは、『高等学校理科 生物基礎/遺伝情報の分配』などの章で説明する。)
DNA量の変化
[編集]配偶子形成の際のDNA量の変化は、原細胞のときを2と置くと、一次母細胞のときは4であり、二次母細胞のときは2となり、卵細胞・精細胞のときは1になり、受精卵のときに2にもどる。 体細胞分裂の際のDNA量の変化は、母細胞のときを2と置くと、前期~終期のときが4であり、娘細胞の時に2にもどる。
DNAと生物の共通性・多様性
[編集]DNAを設計図としタンパク質を作る仕組みは全ての生物で共通している。 しかし、塩基配列が少しずつ変化(ATCGが入れ替わったり、増えたり)して、 生物の多様性が生まれた。
ゲノム
[編集]ゲノム(genome)とはある生物の遺伝子の全体のことである。 2003年にヒトゲノムの解読が完了した。 これにより、ヒトの遺伝子の全体が明らかとなった。 現在では、ゲノム研究は、食品や医療などに応用されている。
(※ ほぼ範囲外:) 遺伝子の本体の研究
[編集]- ※ 2010年代の生物基礎・生物の教科書では、形質転換やファージなどの話題が、あまり見当たらない。
- ※ 数研出版や第一学習社など、いくつかの教科書にあるが、コラム送りになっている。
1869年、スイスのフリードリッヒ・ミーシェルは、 細胞核内の物質を発見しヌクレイン(nuclein)と呼んだ。 当時は、遺伝子の本体はタンパク質であると考えられていたが、 今日では、ヌクレインはDNAと呼ばれ、遺伝子の本体であることが明らかになっている。
- グリフィスの実験

1928年イギリスのフレデリック・グリフィスは、 肺炎レンサ球菌とネズミを用いて実験を行った。 肺炎レンサ球菌には、被膜を持っていて病原性のあるS(smooth)型菌と、被膜が無く病原性のないR(rough)型菌の2種類がある。 被膜の有無と病原性の有無の、どちらも遺伝形質である。 通常の菌の分裂増殖では、S型とR型との違いという遺伝形質は変わらない。
グリフィスの実験結果は次の通り。
- 生きたS型菌をネズミに注射すると、ネズミは肺炎を起こして死ぬ。
- 生きたR型菌をネズミに注射すると、ネズミは肺炎を起こさない。
- 加熱殺菌したS型菌をネズミに注射すると、ネズミは肺炎を起こさない。
- 加熱殺菌したS型菌に生きたR型菌を混ぜてネズミに注射すると、ネズミは肺炎を起こして死ぬ。死んだネズミの血液を調べるとS型菌が繁殖していた。
これはR型菌の形質が、加熱殺菌したS型菌に含まれる物質によって、S型菌の形質へ変化したためであり、
これを形質転換(transformation: nuclein)と呼ぶ。
- アベリーの実験
1943年ころ、カナダのオズワルド・アベリーは、グリフィスの実験での形質転換を起こした物質が何かを特定するため、タンパク質分解酵素とDNA分解酵素を用いて、S型菌・R型菌の実験を行った。
実験結果
- S型菌のタンパク質を分解した抽出液にR型菌を混ぜると、S型菌へ形質転換した。
- 次にS型菌のDNAを分解した抽出液にR型菌を混ぜても、S型菌へ形質転換はしなかった。
これによって、R型菌の形質転換を起こしたのはDNAであることがわかった。
- バクテリオファージの増殖実験

細菌に規制するウイルスのことをバクテリオファージまたは単にファージという。
1952年、アメリカのアルフレッド・ハーシーとマーサ・チェイスは、 T2ファージというファージの一種のウイルスを用いて実験を行った。 T2ファージは細菌に寄生して増殖するウイルスであるバクテリオファージの一種であり、 ほぼタンパク質とDNAからできている。T2ファージの頭部の中にDNAが含まれる。それ以外の外殻(がいかく)はタンパク質で、できている。
彼らは、放射性同位体の35S(硫黄の放射性同位体)および32P(リンの放射性同位体)を目印として用い、硫黄をふくむタンパク質には35Sで目印をつけ、32PでDNAに目印をつけた。DNAは P(リン)をふくむがS(硫黄)をふくまない。彼らの実際の実験では、タンパク質に目印をつけた実験と、DNAに目印をつけた実験とは、それぞれ別に行った。

実験では、それらの放射性同位体をもつT2ファージを大腸菌に感染させ、さらにミキサーで撹拌し、遠心分離器で大腸菌の沈殿と、上澄みに分けた。 大腸菌からは、32Pが多く検出され、あまり35Sは検出されなかった。このことからT2ファージのDNAが大腸菌に進入したと結論付けた。また、上澄みからはT2ファージのタンパク質が確認された。つまり上澄みはT2ファージの外殻をふくんでいる。
さらに、この大腸菌からは、20~30分後、子ファージが出てきた。子ファージには35Sは検出されなかった。
これによって、DNAが遺伝物質であることが証明された。
遺伝子と染色体
[編集]性染色体
[編集]入門的な知識
[編集]- (※ 編集者への注意: ) 2010年代の現代の中学校(高校ではなく)の理科や保健体育では、性染色体を教えてないかもしれません。この節を書く場合、読者が人生で始めて性染色体について習うかもしれない事を念頭に、平易に記述することをお願いします。
- なお、2010年代の現代の中学理科で習うこととして、メンデルの遺伝の法則と、中学生むけに簡略化した減数分裂の理論を習います。
ヒトの体細胞には46個の染色体があり、つまりヒトには23対の染色体がある。(2n=46)
そのうち22対は、男女に共通して存在する染色体であり、これを常染色体(じょうせんしょくたい、Autosome)と呼ぶ。
いっぽう残りの2本の染色体によって、ヒトの性別が決定されるので、これを性染色体と呼ぶ。
ヒトの場合、男女に共通して存在する染色体のことをX染色体という。いっぽう、ヒトでは男性にのみ存在する染色体のことをY染色体という。
- ヒトなどの場合、X染色体があると女性ぽくなり、Yふがあると男性ぽくなるのが、なぜそうだと言い切れるのか、注意ぶかい学生には、気になるかもしれない。なぜなら、YYの組み合わせについては、教科書では何も言及してないからだ。
- つまり、教科書で習った範囲では、まだ「XXやYYだと女性」というYYについての可能性もありうるからだ。
- 答えは実は、通常は性染色体YYの産まれてこないが、まれに産婦から分娩(ぶんべん)される子が性染色体YYの場合があり、子が性染色体YYだと死産になる、という医学的事実がある。(ただし、これは大学の専門課程のレベルの話題であるので、暗記しなくていい。)
- このことから、まずYYの組み合わせについては、高校生は、もう考えなくてよくなる。
- では、次に「X染色体が女、Y染色体が男」ということの分かった理由も学ぼう。答えは、まれに染色体が2対ではなく3対の子が産まれる場合があり、さらに、まれに性染色体が3対でありXXYやXXXやXYYの子が産まれてくる事例がある。XYYのようにY染色体が多いと男性的な特徴がある子が産まれてくることが医学的に分かっている(XYY症候群)。
- いっぽう、XXYの場合の子は、XYと比べると女性的な傾向が比較的に強い(なおこのXXYの人をクラインフェルター症候群という)。
- このほか、XXXの場合、女性的な傾向がある(トリプルエックス症候群。「超女性」ともいう)。XOだと(Oは染色体が無い場合)、女性的である(ターナー症候群)。
- 説明ではいきなり「XXY」とか「XYY」とか、2文字でなく3文字で染色体を表したが、こういうふうに、主に人間において、染色体が2対でなく3対の場合のことをトリソミーという。「トリ」とは、「トリオ」(三人衆)の「トリ」と同様の意味であり、数字の 3 のことである。普通の減数分裂では、トリソミーが起きないが、しかし、まれにトリソミーが起きる場合がある。(※ 生物学では同様の染色体が2対ではなく3対の事例を「三倍体」ともいうが、しかし、どちらかと言うと農作物とかの食料資源で「三倍体」と言う言い回しを使うので、wikibooksでは患者の尊厳に配慮して「トリソミー」という用語を使用した。) なお、トリソミーや三倍体は、性染色体だけでなく常染色体でも起きる場合がある。
- いっぽう、XOのような、ある染色体が1本だけで対にならないの場合、モノソミーという。音響機器などの「モノラル」のモノと同じような意味で、「モノ」は「1個」とかの意味である。
- このように、一般的にヒトの場合、どの性染色体異常でもX染色体の数が多ければ、そのぶん女性的な特徴をもつ。同様に、Yが多ければ、そのぶん男性的な特徴を持つ。ヒトの場合、Y染色体を一つでも性染色体に持つと、男になり、男性器および精巣をもつのが一般的である。(※ これらの話題は大学レベルであり、医療系学部のレベルなので、高校の時点では暗記は不要である。)
- なお、これらは、あくまでヒトの場合であり、ほかの動物の場合は、後述するように、かならずしも、同じような結果になるとは限らない。
参考
[編集]- ※ 教科書には「参考」(数研出版の教科書)としてコラム的にページ下部に書かれている。啓林館の教科書では、本文中に書かれている。
ヒト以外の動物も含めると、性の決定には、XY型、XO型、ZW型、ZO型の4つがある。 XY型は、雌が同形のXX、雄が異形のXYの性染色体をもち、 ショウジョウバエや、ヒトなどの哺乳類が行う。 XO型は、雌が同形のXX、雄がXの1つだけの性染色体をもち、 トンボやバッタなどが行う。 ZW型は、雌が異形のZW、雄が同形のZZの染色体をもち、 ニワトリ、ヘビ、カイコガなどが行う。 ZO型は、雌がZの1つだけ、雄が同形のZZの染色体をもち、 スグリエダシャクなどが行う。
- ※ 範囲外: (※ 検定教科書(生物基礎・生物)に記述なし)
- ただし、性の決定を性染色体以外で行う生物も多くいる。
- 例えば、アカウミガメは、卵が孵化する際の温度で雌雄が決まる。
- また、カタツムリは、雌雄同体で、一つの個体が精巣と卵巣を持つ。
遺伝の法則
[編集]遺伝と形質
[編集]遺伝(heredity)とは、生物の形や性質が、遺伝子によって、親から子へ伝わることである。
また、生物の形や性質のことを形質(けいしつ、trait)と呼ぶ。 形質には親から子へ遺伝する遺伝形質(genetic trait)と、 環境の影響によって獲得した遺伝しない獲得形質(Acquired trait)がある。 このページでは、形質とは遺伝形質を指す。
生殖の際に、親から生殖細胞を経て、子に伝えられている遺伝の因子を遺伝子(いでんし)といい、こんにちでは遺伝子の正体は、細胞にふくまれるDNA(ディーエヌエー)という物質であることが知られている。
メンデルの法則
[編集]- ※ 2010年代の現代の中学(高校ではなく)で、「メンデルの法則」「優性」「劣性」という用語を習ってある。これ以外の用語は、中学の遺伝の単元では習わないようだ。
- また、2010年代の高校カリキュラムでは、下記の内容は高校3年の専門『生物』(かつての『生物II』に相当)に移動している。
メンデルの実験
[編集]メンデルは、異なる形質をもつエンドウの品種を用意し、2年間にわたり育て、 同一個体の配偶子間で行われる自家受精(autogamy)で 全く同じで変化しない子孫を生じる純系(pure line)の品種を選んだ。 その際、明確に決定的に発現する、互いに異なる対立形質を7つ採用し、 1856年から62年にかけて交配実験を行った。
- 熟した種子の形の違い(丸・しわ)
- 種子の胚乳の色の違い(黄・緑)
- 種皮の色の違い(有色・無色)
- 熟したさやの形の違い(ふくれ・くびれ)
- 未熟なさやの色の違い(緑・黄)
- 花の位置の違い(腋生(えきせい)・頂生(ちょうせい))
- 茎の長さの違い(高い・低い)
実験1 1.種子の形について、 丸としわの純系を用意して両親P(Parents)としたところ、 その子雑種第一代[2]F1(Filius)は、全て丸であった。 このようにF1では、対立形質の片方のみが表れる。 現れる形質を優性形質(dominant trait)と呼び、現れない形質を劣性形質(recessive trait)と呼ぶ。 ここでの優性・劣性は、単に形質が現れやすい・現れにくいを意味し、形質が優秀である・劣等であるを意味しない。
実験2 F1を自家受精したところ、 雑種第二代F2では丸としわが5474個と1850個で、およそ3:1の出現比であった。 このようにF2では、 優性形質と劣性形質がおよそ3:1の比で出現する。
実験3 F2を自家受精したところ、 F2でしわだったものは、F3で全てしわの純系となり、 F2で丸だったものは、565株のF3の内、 193株は丸の純系となり、 372株は丸としわを3:1の比で生じた。 このようにF3では、F2で劣性形質を示すものは、劣性形質の純系となり、 F2で優性形質を示すものは、このうち、3分の2は優性形質と劣性形質を3:1の比で生ずる子孫を作り、 3分の1は優性形質の純系となる。
実験4 1.種子の形と2.胚乳の色について、 種子の形が丸で胚乳の色が黄の純系と種子の形がしわで胚乳の色が緑の純系を用意して両親Pとしたところ、 その子F1はすべて丸で黄であった。
実験5 F1を自家受精したところ、 F2では丸・黄、丸・緑、しわ・黄、しわ・緑が315個、108個、101個、32個で、 およそ9:3:3:1の出現比であった。
遺伝子型と表現型
[編集]個体の遺伝子の構成を記号で表したものを遺伝子型(genotype)と呼ぶ。 遺伝子型はふつう優性形質をアルファベットの大文字で表し、 劣性形質をアルファベットの小文字で表す。 ある形質を決定する遺伝子は、 ペアの染色体の同じ位置に1つずつ、 あわせて2つあるため、 アルファベット2文字で表す。(例:AA,Aa,aa)
また、AAやaaのように同じ遺伝子がペアになっているものをホモ接合体(homozygous, 同型接合体)と呼び、 Aaのように異なる遺伝子がペアになっているものをヘテロ接合体(heterozygous, 異型接合体)と呼ぶ。
遺伝子型によって現れる形質を表現型(phenotype)と呼ぶ。 遺伝子型の記号を[]で囲んで表すこともある。(例:[A],[a])
検定交雑と戻し交雑
[編集]遺伝子型の判別のために、その個体と劣性形質の個体とを交雑することを検定交雑(test cross)と呼ぶ。 また、F1とPとを交雑することを戻し交雑(backcross)と呼ぶ。 下の表は、検定交雑で遺伝子型を判別する方法を示している。 配偶子?2と?4の遺伝子構成は、F1の表現型とその分離比から予想できる。 つまり、?2はAのみ、?4はAとaである。 両親?1と?3の遺伝子型は、配偶子?2と?4の遺伝子構成から予想できる。 つまり、?1はAA、?4はAaである。
| P | 優性形質 ?1 |
劣性形質 aa | |
| ↓ | ↓ | ||
| 配偶子 | ?2 | 丅 | a |
| ↓ | |||
| F1 | 全て優性形質 Aa |
| P | 優性形質 ?3 |
劣性形質 aa | |
| ↓ | ↓ | ||
| 配偶子 | ?4 | 丅 | a |
| ↓ | |||
| F1 | 優性形質:劣性形質 Aa:aa 1:1 |
優性の法則
[編集]| Pの配偶子 | ||
|---|---|---|
| A | ||
| Pの配偶子 | a | Aa [A] |
実験1では、 種子の形が丸をA,しわをaと表すとすると、 遺伝子型は、丸の純系はAA、しわの純系はaaと表せる。 この両親Pの配偶子はそれぞれA、aとなり、 その子F1の遺伝子型はAaとなり、表現型は[A]となる。 このように、優性形質の純系と劣性形質の純系とを交雑すると、 その子は優性形質のみを表し、 これを優性の法則(law of dominance)と呼ぶ。 なお、今日では、エンドウの種子の形を決める遺伝子は、 実際には酵素を作る遺伝子であり、その酵素がデンプンを作って種子の形を丸にしていることがわかっている。デンプンの量は、AaはAAとaaの中間であるが、種子の形を丸にするには十分な量であるため、Aaの種子の形は丸となっている。
分離の法則
[編集]| F1の配偶子 | |||
| A | a | ||
| F1の配偶子 | A | AA [A] |
Aa [A] |
| a | Aa [A] |
aa [a] | |
実験2では、 F1の遺伝子型はAaと表され、 配偶子が作られるとき分離し、 それぞれの配偶子はA,aとなる。 このように配偶子形成の際ペアの遺伝子が分離し、 それぞれ配偶子に受け継がれることを分離の法則(law of segregation)と呼ぶ。 F1の自家受精では、 その配偶子がそれぞれ受精するため、 F2ではAA:Aa:aa=1:2:1となり、 結果[A]:[a]=3:1となる。
実験3では、 F2で[a]だったものは、aaであるから、 その配偶子はaであり、自家受精でaaつまり[a]となる。 F2で[A]だったものは、AA:Aa=1:2であるから、 3分の1のAAの配偶子はAであり、自家受精でAAつまり[A]となり、 3分の2のAaの配偶子はA,aとなり、自家受精でAA:Aa:aa=1:2:1つまり[A]:[a]=3:1となる。
独立の法則
[編集]| Pの配偶子 | ||
|---|---|---|
| AB | ||
| Pの配偶子 | ab | AaBb [AB] |
実験4では、 種子の形が丸をA,しわをa、胚乳の色が黄をB,緑をbと表すとすると、 遺伝子型は、丸で黄の純系はAABB、しわで緑の純系はaabbと表せる。 この両親Pの配偶子はそれぞれAB,abとなり、 その子F1の遺伝子型はAaBbとなり、表現型は[AB]となる。
| F1の配偶子 | |||||
| AB | Ab | aB | ab | ||
| F1の配偶子 | AB | AABB [AB] |
AABb [AB] |
AaBB [AB] |
AaBb [AB] |
| Ab | AABb [AB] |
AAbb [Ab] |
AaBb [AB] |
Aabb [Ab] | |
| aB | AaBB [AB] |
AaBb [AB] |
aaBB [aB] |
aaBb [aB] | |
| ab | AaBb [AB] |
Aabb [Ab] |
aaBb [aB] |
aabb [ab] | |
実験5では、 F1の遺伝子型はAaBbとあらわされ、 配偶子が作られるとき分離し、 それぞれの配偶子は、AB,Ab,aB,abとなる。 F1の自家受精では、 その配偶子がそれぞれ受精するため、 F2でAABB:AABb:AaBB:AaBb:AAbb:Aabb:aaBB:aaBb:aabb=1:2:2:4:1:2:1:2:1となり、 結果[AB]:[Ab]:[aB]:[ab]=9:3:3:1となる。
実験4・5では、 種子の形だけあるいは胚乳の色だけに注目すると、 それぞれ優性の法則と分離の法則に従い独立して遺伝している。 つまり、種子の形に関しては[A]:[a]=3:1であり、胚乳の色に関しては[B]:[b]=3:1である。 このように、2つの遺伝子が異なる染色体に存在するとき、 その遺伝子が互いに影響しないことを独立の法則(law of independence)と呼ぶ。
参考: さまざまな遺伝
[編集]- ※ この節の話題は、かつ2000年代ごろまで、下記の不完全優性~抑制遺伝子、伴性遺伝などの話題は、むかしは高校生物の教科書や参考書に良くある話題だったが、しかし現代の高校教育では重要度が低いと考えられるように教育状況が変化しており(『もういちど読む』シリーズの高校生物にその事情が書いてある)、検定教科書では「参考」などのコラムに送られている。
不完全優性
[編集]優性と劣性の関係が不完全な遺伝の仕方を不完全優性(incomplete dominance)と呼ぶ。 不完全優性では優性の法則は当てはまらない。
不完全優性は、マルバアサガオなどが行う。 マルバアサガオには、花の色が赤Rと白rのものがある。 花の色が赤の純系RRと白の純系rrを両親Pとすると、 その子F1はRrで花の色が中間の桃色となる。 さらにその子F2は、RR:Rr:rr=1:2:1で、赤色:桃色:白色=1:2:1となる。
| P | RR 赤色 |
× | rr 白色 |
| ↓ | |||
| F1 | Rr 桃色 |
||
| ↓ | |||
| F2 | RR:Rr:Rr:rr 赤色:桃色:桃色:白色 | ||
致死遺伝子
[編集]成体になるまでに致死作用がある遺伝子を致死遺伝子(lethal gene)と呼ぶ。
致死遺伝子は、多くの生物に存在する。 例えば、ハツカネズミは致死遺伝子を持っており、 毛の色が黄色Yと灰色yのものがある。 黄色Yyを両親Pとすると、 その子F1はYy:yy=2:1で、[Y]:[y]=2:1となる。 YYの個体は発生の段階で死んでしまう。 これはYが劣性の致死遺伝子だからである。
| P | Yy 黄色 |
× | Yy 黄色 |
| ↓ | |||
| F1 | YY:Yy:Yy:yy 死:黄色:黄色:灰色 | ||
複対立遺伝子
[編集]同一の遺伝子座にある、同一形質を決める、複数の遺伝子を複対立遺伝子(multiallelic gene)と呼ぶ。
複対立遺伝子には、ヒトのABO式血液型などがある。 ヒトのABO式血液型には、A型、B型、AB型、O型の4種類があり、 AとBとは不完全優性で、A,BはOに対して完全優性である。 例えば下の表のように、AO(A型)とBO(B型)を両親とすると、 その子はAB,AO,BO,OOとなり、それぞれAB型,A型,B型,O型となる。
| 表現型 | A型 | B型 | AB型 | O型 |
|---|---|---|---|---|
| 遺伝子型 | AA AO |
BB BO |
AB | OO |
| P | AO A型 |
× | BO B型 |
| ↓ | |||
| F1 | AB:AO:BO:OO AB型:A型:B型:O型 | ||
補足遺伝子
[編集]対立しない2つ以上の遺伝子が、その働きを互いに補足しあって1つの形質を決めるとき、その遺伝子を補足遺伝子()と呼ぶ。
補足遺伝子には、スイートピーの花の色などがある。 色素原を作る遺伝子をC、色素原から色素を作る遺伝子をPとし、 白色花CCppと白色花ccPPを両親Pとすると、 その子F1はCcPpで有色花となる。 さらにその子F2は、C-P-:C-pp:ccP-:ccpp=9:3:3:1で、有色花:白色花:白色花:白色花=9:3:3:1つまり有色花:白色花=9:7となる。 これはCとPの両方をもっていないと色素が作られないためである。
| P | CCpp 白色花 |
× | ccPP 白色花 |
| ↓ | |||
| F1 | CcPp 有色花 |
||
| ↓ | |||
| F2 | 9C-P-:3C-pp:3ccP-:1ccpp 9有色花:3白色花:3白色花:1白色花 | ||
抑制遺伝子
[編集]他の遺伝子の働きを抑制する遺伝子を抑制遺伝子(suppressor gene)と呼ぶ。
抑制遺伝子には、カイコガのまゆの色などがある。 黄色遺伝子をY、Yの働きを抑制する遺伝子をIとし、 白まゆIIyyと黄まゆiiYYを両親Pとすると、 その子F1はIiYyで白まゆとなる。 さらにその子F2は、I-Y-:I-yy:iiY-:iiyy=9:3:3:1で、白まゆ:白まゆ:黄まゆ:白まゆ=9:3:3:1つまり白まゆ:黄まゆ=13:3となる。 これはIがYの働きを抑制するためである。
| P | IIyy 白まゆ |
× | iiYY 黄まゆ |
| ↓ | |||
| F1 | IiYy 白まゆ |
||
| ↓ | |||
| F2 | 9I-Y-:3I-yy:3iiY-:1iiyy 9白まゆ:3白まゆ:3黄まゆ:1白まゆ | ||
伴性遺伝
[編集]性染色体の中にあるが、性の決定以外の働きをもった遺伝子の遺伝現象のことを伴性遺伝(sex-linked inheritance)という。 伴性遺伝は形質の発現が性別によって異なり、 ヒトの赤緑色覚異常や血友病などに見られる。
ヒトの赤緑色覚異常の遺伝子は、X染色体上にある劣性遺伝子である。
- 男性はそもそもX染色体を一つしかもたないので、そのX染色体に色覚異常遺伝子があれば、発症する。
- いっぽう女性は、X染色体を2つもつので、両方のX染色体に色覚異常遺伝子がある場合にだけ発症する。女性において線染色体の片方だけに色覚異常の遺伝子がある場合、発症はしない。(※ 範囲外: しかし保因者(ほいんしゃ)である。)
記号的に書けば、優性遺伝子と劣性遺伝子をそれぞれA,aと表すと、 XA、Xaのように表す。この場合、Xaが色覚異常の遺伝子である。
- 男性では、XAY、XaYの2種類の遺伝子型があり、XaYの場合のみ色覚異常となる。
- 女性では、XAXA、XAXa、XaXaの3種類の遺伝子型があり、XaXaの場合のみ色覚異常となる。
このように、伴性遺伝は性別によって遺伝の仕方が異なる。
このようなメンデル遺伝的な理由もあって男性のほうが統計的には遺伝性の色覚異常は多いが、しかし女性でも遺伝性の色覚異常者はいる[3]。
- ※ しばしば、「色覚異常は男性だけ」のような誤解の書かれている医療入門書もあるが、そのような入門書の記述は間違いである。
連鎖と組み換え
[編集]同じ染色体にある遺伝子が、配偶子形成の際に行動をともにすることを、遺伝子の連鎖(linkage)という。 1905年、イギリスのウィリアム・ベーツソンは、 スイートピーの交雑実験から、 連鎖の現象を発見した。
生殖細胞の減数分裂のとき、相同染色体の一部が交換する現象を乗換え(crossover)という。 そのときに遺伝子の配列が変わることを組換え(Recombination)という。 遺伝子の組換えが起こる割合を組換え価()といい、パーセントで表される。
組み換え価を %、組み換えの起こった配偶子数を、全ての配偶子数をと置くと、組み換え価は次のように求める。






1926年、アメリカのトーマス・ハント・モーガンは、 異なる3つの形質に対し、組み換え価を求め、その組み換え価から遺伝子距離を求める三点交雑(three-point cross)により、 キイロショウジョウバエの遺伝子の配列を図示し、これを染色体地図(chromosome map)と呼ぶ。
脚注
[編集]- ^ 岩槻邦夫、須原準平訳『メンデル 雑種植物の研究』岩波書店、1999年発行
- ^ メンデルは「雑種第一代」を単に「雑種」と呼び、「雑種第二代」を「雑種第一代」と呼んでいた。
- ^ KIM E. BARRETT ほか原著改訂、岡田泰伸 監訳『ギャノング生理学 原著23版 』丸善株式会社、平成23年1月31日 発行、P233、節『クリニカルボックス 12-6 色覚異常』
参考文献
[編集]- 田中隆荘ほか『高等学校生物I』第一学習社、2004年2月10日発行、pp.110-154
- 『NHK高校講座 生物』第16-21回
- 生物学用語辞典 - Weblio 学問